
SYSTEMATIC REVIEW
Varicella (Human Herpes Virus-3) Vaccine Potential Role Against Herpes (HSV-1/HSV-2) Viruses to Prevent HIV-1 Pandemic in Sub- Saharan Africa
Jacqueline Le Goaster1, *, Patrice Bourée1, Franck N. El Sissy1, Johanna Pokossy Epee1, Frédéric Tangy2, Anne-Lise Haenni3, Allan Goldstein4
Article Information

Identifiers and Pagination:
Year: 2018Volume: 10
First Page: 116
Last Page: 123
Publisher ID: TOIDJ-10-116
DOI: 10.2174/1874279301810010116
Article History:
Received Date: 31/5/2018Revision Received Date: 27/7/2018
Acceptance Date: 6/8/2018
Electronic publication date: 31/08/2018
Collection year: 2018

open-access license: This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International Public License (CC-BY 4.0), a copy of which is available at: https://creativecommons.org/licenses/by/4.0/legalcode. This license permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Background:
Synergy exists between DNA and RNA viruses. It was found that the Human Immunodeficiency Viruses (HIV-1) are RNA viruses at the origin of Acquired Immune Deficiency Syndrome (AIDS). The DNA recurrent herpes diseases are associated to AIDS virus at the origin of Sub-Saharan cancer AIDS pandemic.
Objective:
It is speculated that a varicella virus (HHV-3) immune defect could originate HSV- 1/HSV-2 recurrent herpes diseases that can be cured by varicella vaccine (2012).
Methods:
At a Symposium held in Kampala, Uganda (1962), impressive Sub-Saharan cancer epidemics: Hodgkin lymphomas and Kaposi sarcomas have been reported since the onset of the 20th century and remained unexplained. Over one thousand publications related to these cancer epidemics were presented. For millenniums, Bantu populations have been living in tropical forests close to chimpanzees infected by Simian Immune Deficiency viruses (SIV). SIV became Human Immune Deficiency viruses (HIV-1). AIDS is a zoonosis.
Results:
The DNA and RNA viruses, herpes with HIV-1 viruses, are correlated to Sub- Saharan AIDS infections. They induce an extensive immune deficiency with other herpes viruses such as HHV-4 and HHV-8, which are linked to lymphomas and Kaposi sarcomas. It is postulated that a primary HHV-3 immune weakness could be linked to herpes partnership with AIDS pandemic.
Conclusion:
The Oka, anti-HHV-3, varicella vaccine is able to cure HSV1/HSV2 recurrent herpes diseases. It induces a specific increase of the varicella antibodies. Thus varicella vaccination could prevent herpes recurrences in Sub-Saharan Africa. One- child dose varicella vaccine could be proposed as the first step to overcome HHV-3 herpes deficiency in order to prevent AIDS pandemic.
1. INTRODUCTION
For more than a century, unexplained cancers such as Hodgkin lymphomas and Kaposi sarcoma have occurred in Subtropical Africa (Fig. 1).
 |
Fig. (1).
Correlation between Kaposi’s sarcoma and Bantu ethnies in subtropical Africa. Location of Bantu populations. Location of Bantu populations.  Sites of medical centers, in tropical Africa (1914-1960). Sites of medical centers, in tropical Africa (1914-1960).  Chimpanzee observatories (1914-1960). Chimpanzee observatories (1914-1960).
|
In 1981 in San Francisco, this same type of tumors was related to unidentified viral infections; in 1983, HIV-1 was identified as being at the origin of AIDS cancer pandemic. Later in 2000, it was discovered that herpes viruses have a pathogenic power at the origin of tumors such as Hodgkin lymphomas, Kaposi sarcomas, etc.
It was established that these herpes viruses play a key role related to HIV-1 infections [1].
The human herpes simplex viruses HSV-1/HSV-2 and the varicella zoster virus HHV-3 are members of the subfamily Alphaherpesvirinae following the morphological order of herpes viruses distinct from all other viruses [2]. (Fig. 2).
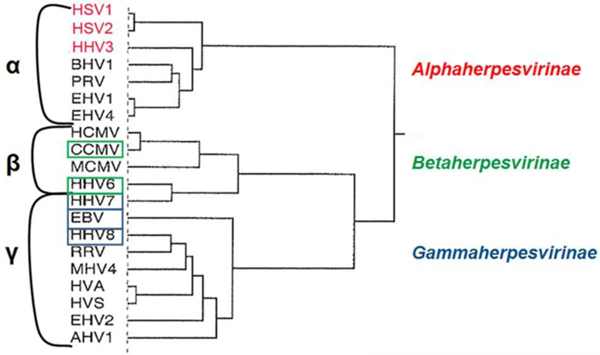 |
Fig. (2). Phylogenetic tree of the members of the Herpesviridae family [2]. |
The members of the Herpesviridae phylogenic tree refer to herpes virus pathogeny for humans: α herpesvirinae, β herpesvirinae and γ herpesvirinae according to McGeoch [3] (Fig. 3).
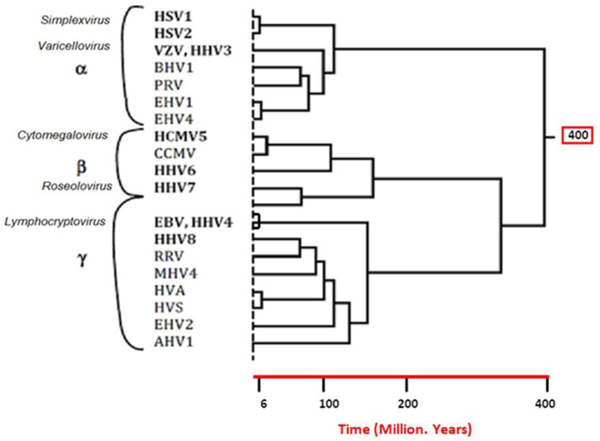 |
Fig.(3). Phylogenetic tree of the Herpesviridae family includes numerous living animals [2]. |
Infections caused by HSV-1 or HSV-2 result of cell-to-cell transmission, involve cell-mediated immunity [4]. The HHV-3/HSV-1/HSV-2 viruses must have a close relationship, that has however been sparsely explored by researchers in fundamental virology [5, 6].
“Varicella is a common and very infectious disease in children” caused by VZV, known as varicella zoster virus (HHV-3) [7]. “The causative agent of varicella is one of the eight herpes viruses that include humans as natural hosts. “The Varicella Zoster Virus exhibits a low genomic diversity relatively to other studied herpesviruses” [8].
The varicella zoster virus (VZV/HHV-3) can be considered as a primordial event with a specific immunogenic Human Leucocyte Antigen (HLA) as a final result.
It is speculated that a varicella virus (HHV-3) immune defect could originate HSV- 1/HSV-2 recurrent herpes diseases which can be cured by varicella vaccine. Since 2005, based on this hypothesis, the varicella vaccine is used against herpes viruses, it has been efficient. It is possible to acquire positive long-term clinical results as it has been observed [9]. There have been no relapses concerning more than 70 patients (10 to 80 years old) so far.
The HSV-1/HSV-2 recurrent herpes diseases play a key role related to HIV-1 infections. The varicella vaccine will be used to cure herpes diseases and it could play a role, first against HSV-1/HSV-2 recurrent herpes diseases, further HIV-1 infections [9].
The first HHV-3 vaccine was the “Oka strain” vaccine, created by Takahashi et al. (1974) [7]. The original virus strain was used initially to prevent varicella in high-risk leukemic children who remained free from varicella with safety.
The varicella vaccine is a live attenuated herpes virus vaccine (1,350 UFP), available for children and adults. In the USA, in 1995, varicella vaccination program was decided using: an universal one dose childhood. It became an obligation for all children in 3 states: California, Massachusetts, Pennsylvania. The incidence of Varicella has fallen by 80%.
Studies of geographical variations of the varicella virus (VZV/HHV-3) have been performed. Isolates from Caucasian populations are represented by just one genome sequence. The strain related to the OKA vaccine is constituted by Japanese origin and an incomplete VZV Sub-Saharan African genome sequence [2].
Therefore the OKA varicella vaccine is originated partly from Japan and partly from Sub-Sahara. This underlines the ancient human origin from sub-tropical Africa. This varicella vaccine is used to prevent shingles and zoster in immunodepressed individuals. The varicella vaccine has potential pharmacodynamic properties but these have not been deeply investigated [8]. However, recently the efficiency of such anti-VZV vaccine against recurrent HSV-1/HSV-2 infections has been already reported [9].
It appeared essential to increase the efficient HHV-3 (VZV) power to raise the immungenetic ability against recurrent HSV-1/HSV-2 herpes diseases. Then, as it is reported it is possible to prevent recurrent herpes diseases linked to HIV-1 infections [1, 9, 10].
The effect of the varicella vaccine on recurrent herpes diseases must be attributed to a cell-mediated immune response as it has been reported [9, 11]. It suggests immunological induction of cell-mediated reactivity between HSV-1/HSV-2 and HHV-3 revealed by a specific raise of anti-HHV-3 Ab. induced by Varicella Vaccine.
Lymphocyte stimulation triggers the observed clinical anti-HSV-1/anti-HSV-2 efficiency using the varicella vaccine to prevent recurrent herpes diseases. It is clearly in favor of this hypothesis, a cell-mediated reactivity might be involved between HSV- 1/HSV-2 and HHV-3. As a matter of fact, a specific serological increase of anti-HHV-3 Ab levels correlated with the recurrent herpes diseases cure can be observed.
In 2000, a link was established between the sexually recurrent herpes disease HSV-1/HSV-2 with HIV-1 AIDS diseases which were prevalent in Subtropical Africa [12].
It is important to underline the role played by the other members of the Betaherpesvirinae as HHV-5, the other members of the Gammaherpesvirinae include HHV-4 known as Epstein-Barr viruses are involved in lymphomas including Hodgkin lymphomas. Finally the HHV-8 viruses are linked to Kaposi sarcomas (Table 1).
| HSV-1 | Herpes Simplex Virus Type-1 (oro-buccal herpes) |
| HSV-2 | Herpes Simplex Virus Type-2 (genital herpes) |
| HHV-3 VZV | Varicella Zoster Virus, Human Herpes Virus-3. |
| HHV- 4 EBV | Human Herpes virus-4, Epstein-Barr Virus (mononucleosis, Hodgkin’s lymphoma). |
| HHV- 5 CMV | Human Herpes Virus-5, Cytomegalovirus (spontaneous abortion). |
| HHV- 6 | Human Herpes Virus-6 (Roseolovirus……..) |
| HHV- 8 | Human Herpes Virus-8 (Kaposi sarcoma) |
2. METHODS – DATA ANALYSIS
Subtropical African Cancer Epidemics in Bantu populations were reported in the early 20th century as an epicentre of Kaposi sarcomas and lymphoma epidemics.
Many clinicians in Central Africa proposed an infectious viral origin for outdoor Bantu tribes workers (1914). They failed to prove it. A resurgence of interest was stimulated by the high incidence of these cancers observed in South African histological samples tested in Johannesburg [13, 14]. Clinicians observed that outdoor Bantu hunters were the selected targets of these cancer epidemics [15]. They were exposed to the blood of chimpanzees which could be infected by Simian Immunodeficiency Virus (SIV). AIDS IS A ZOONOSIS [16]. Primates are reservoirs for SIV and if they carry SIV they can contaminate the Bantu hunters, SIV thus became HIV-1 in human-beings. SIV and HIV-1 are both RNA viruses [16]. Central Africa remains the global HIV-1 epicentre (Fig. 1).
HIV-1 viruses escape from immune and drug pressures. It appears to be a co-activator of micro-organisms such as Pneumocystis jiroveci and viruses such as herpes viruses: lymphomas and Kaposi sarcomas as it was observed in the San Francisco epidemic (1981) [17]. In 2008 close similarities were observed between the African Kaposi sarcomas epidemics and the same epidemics in San Francisco. Mostly young adults were affected by these similar fatal diseases called Acquired Immune Deficiency Syndrome, AIDS disease, which then emerged worldwide.
The production of deficient anti-HHV-3 antibodies is postulated in front of recurrent herpes viruses HSV- 1/HSV-2, this could enhance the herpes immune deficiency facing HIV-1.
All human beings suffer from shingles caused by varicella virus (HHV-3/VZV) during childhood but an immunogenetic HHV- 3 failure can be a first prerequisite to favour HSV-1/HSV-2 recurrent herpes diseases followed by the fatal outcome HIV-1 cell anchorage as stated by Lawrence Corey [18].
3. RESULTS
The evaluation of cancer epidemic data showed, Specific Environmental Factors that were proposed to play a key role in cancer epidemics in Subtropical Africa [19]. These cancers are most frequent in South Africa as well as East Africa where the main part of the Bantu population is composed of outdoor forest farmers and where bare footedness is the rule. This suggested an etiological role of chronic injury among Bantu hunter inhabitants of tropical forests. Surprisingly, no case was reported among booted Bantu clerks living in cities in South Africa [19] (Figs. 4 and 5).
 |
Fig. (4). Kaposi sarcomas and lymphoma epidemics (1914-1960) in [16] Among black African populations report by Findlay [28]. |
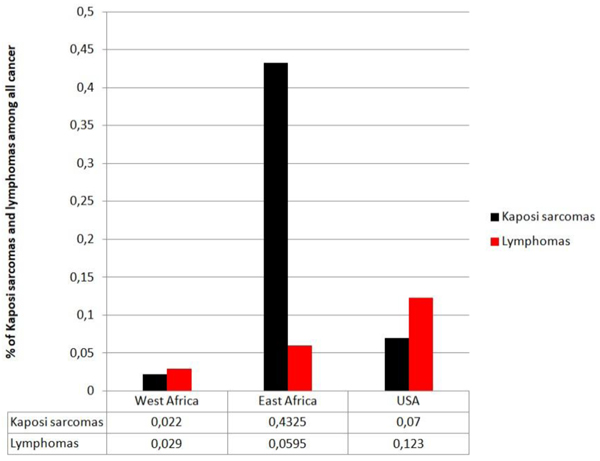 |
Fig. (5). Kaposi’s sarcoma and lymphoma epidemics among all cancers from 1988 to 1997 in Gambia. Compared to the USA these cancers were related to AIDS diseases in Bah et al. [27]. |
In Tropical Africa, Whites and Indians remained relatively unaffected by Kaposi sarcomas and lymphomas. We can wonder why they were able to save their lives compared to the native tropical forest Bantu populations?
In industrialized countries such as in Europe and in the USA, Kaposi sarcomas are known as a disease of the elderly compared to the large infected young Bantu populations (Fig. 6).
 |
Fig. (6). Symposium in Kampala, Uganda, on Kaposi sarcomas, lymphomas and all cancers in Africa (1914-1960), series in black populations, Findlay [28]. |
These Kaposi sarcomas and lymphomas were already proposed to be contagious [20, 21] with an incubation period of 1 to 10 years respectively in Mozambique [22] linked to occupations, occurring among farmers who constitute 95% of the Bantu population in Cameroon, Gabon, Congo [13].
Kaposi sarcomas identified at the histological level appeared at that time as clinically unique compared to American or European Kaposi sarcomas in terms of symptoms and fatal evolution [23].
4. DISCUSSION
Subtropical African human populations living in tropical forests were contaminated by zoonotic or cross-species transmission. Monkeys represent the natural reservoir of simian viruses, their infected females by SIV breast-fed for 4 years, so their simian babies were infected by SIV and these orphan animal were kept as pets which are able to contaminate all the hunter families [24, 25].
For over several centuries, there have been frequent opportunities for human beings to become infected by SIV-infected chimpanzees but it is important to note that genetic mechanisms have been operating to enhance the immunogenic power of human beings to escape and survive attacks by endemic HIV-1 strains in Subtropical Africa.
It is necessary to help people in their immunological power against HIV-1. It is the medical priority of all physicians, as Lawrence Corey wrote it: the “two unrelated pathogens,HSV- 1/HSV-2 and HIV-1, occupy the same ecological space in Central Africa” [18]. The observations reported here underline the basic role played by members of the Herpesviridae family at the onset of the HIV-1 pandemic worldwide [26].
Bantu simian hunters could have presented an immungenetic weakness against VZV with an another reason: consanguinity. They became the deficient victims of the HSV-1/HSV-2 recurrent herpes diseases. Then exposed to be infected by SIV, the chimpanzee hunters and their families became eligible for SIV/HIV-1 contaminations and AIDS disease.
CONCLUSION
Following the recent discovery of the links between HSV-1/HSV-2 recurrent herpes diseases with a first varicella (HHV-3) immune deficiency, it appears that there is a preliminary biological herpes viral defect able to precede and to favor HIV- 1 infection.
It is logical to postulate a preventive anti-herpes therapy using varicella vaccine. The use of a universal child dose of varicella vaccine could be the primary objective to cure the recurrent herpes infections and then to prevent AIDS pandemic.
This will represent the first step to push back herpes diseases related to HIV-1/AIDS pandemic.
Moreover, a first HHV-3 failure can precede or induce a large immune deficiency against all the other pathogenic herpes viruses such as HHV-4, HHV-5, HHV-6 and HHV-8.
Therefore, the use of varicella vaccine could represent the hope to prevent HSV1-/HSV-2 infections related to AIDS pandemic.
FUNDINGS
The authors thanked Physiologie et Tumeur, an independant non-profit Research Institution in Paris, France, for its support.
PATIENT AND PUBLIC INVOLVEMENT STATEMENT
Starting at the early 20th century, the authors have analyzed African population’s historical data. No recent patients were involved or concerned in this study.
CONSENT FOR PUBLICATION
Not applicable.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
The authors thank Professor Allan Goldstein for his constructive comments and valuable suggestions regarding the manuscript and Professor Michiaki Takahashi †, who originally created the Oka strain anti-varicella vaccine, for his enthusiasm, constant support, and encouragement to publish our research.





